Предметом изучения генетики являются два неразрывных свойства всех живых организмов – наследственность и изменчивость. Изменчивость представлена многообразием форм внутри каждого вида, породы и даже одного помета. Но в то же время все представители одного вида и одной породы имеют несомненное сходство обеспечиваемое наследственностью.
Для каждого вида животных характерен набор хромосом с определенными характеристиками (число, форма, размер и т.д.), составляющий кариотип.
Половые клетки содержат одинарный набор хромосом, называемый гаплоидным, который принято обозначать одной буквой n. Клетка, образованная путем слияния двух половых клеток, содержит в два раза большее количество хромосом, так называемый, диплоидный набор – 2n. Каждая пара хромосом диплоидного набора представлена гомологичными хромосомами, одна из которых получена от отца, а другая от матери. Диплоидный набор собаки содержит 78 хромосом.
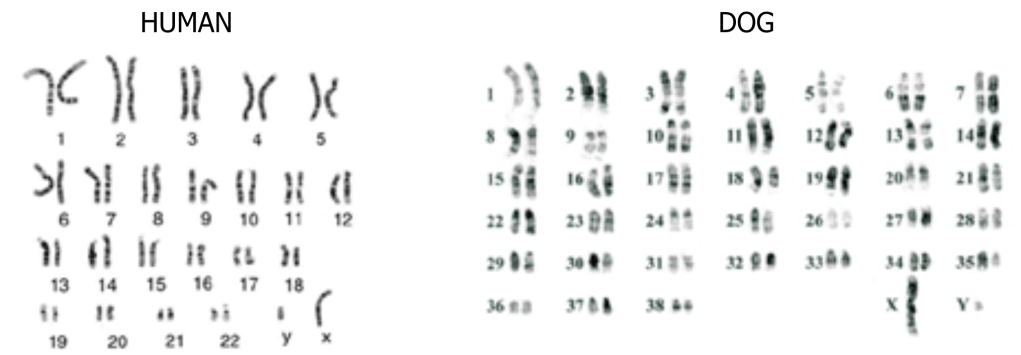
Несмотря на разное количество хромосом, геном собаки содержит приблизительно
столько же генов, сколько и геном человека, около 20 000.
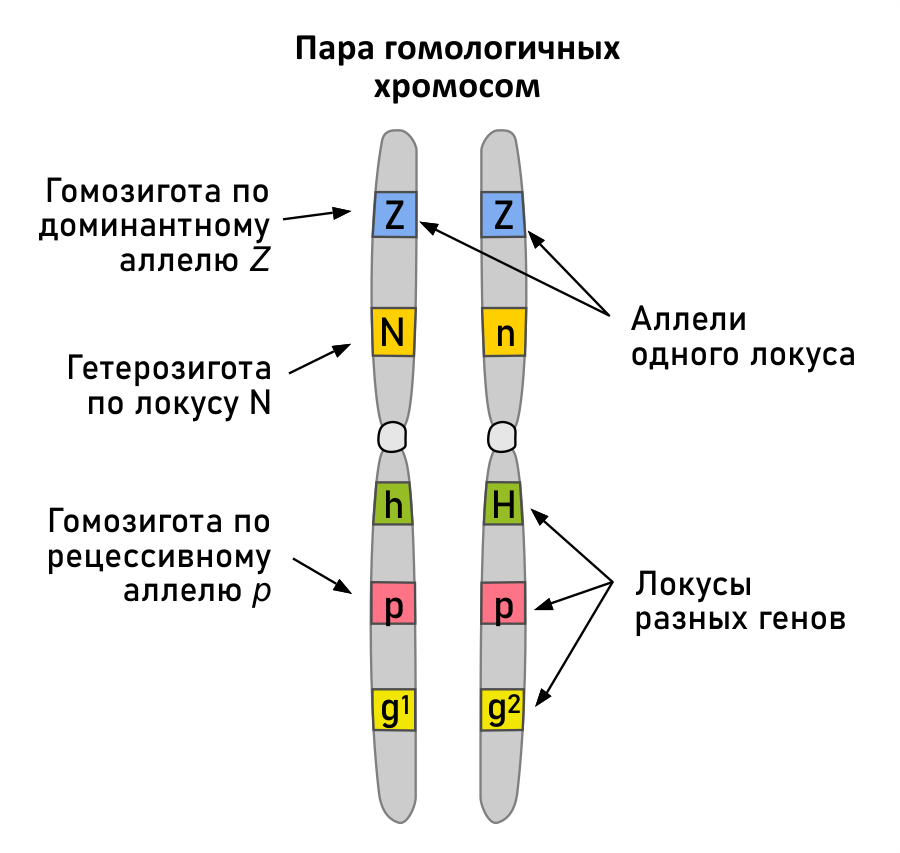
Все наследственные свойства и признаки определяются вполне определенными независимыми друг от друга материальными единицами – генами. Каждый ген занимает строго определенное место в строго определенной хромосоме, называемое локусом. В силу парности хромосом в клетках, гены в хромосомном наборе также представлены парами. Гены могут претерпевать изменения – мутировать, вследствие этого могут изменяться и внешние проявления признака, за который отвечает данный ген. Различные формы одного и того же гена называют аллелями. Понятия проиллюстрированы ниже на примере локуса D.

Не каждая мутация приводит к изменению фенотипа. Мутации могут быть нейтральными или незначительно изменяющими работу гена.
Особи, получившие от отца и матери одинаковые аллели одного гена, называются гомозиготными, а разные – гетерозиготными по данному признаку. Гены и локусы принято обозначать буквами латинского алфавита, например, локусы А, E, гены ASIP, MC1R и т.п.
Доминирование
Находясь в гетерозиготном состоянии, аллели определенным образом взаимодействуют друг с другом. Случай, когда один из них полностью подавляет действие другого, называется полным доминированием. Доминантный аллель обычно обозначается заглавной буквой латинского алфавита, рецессивный — строчной. При полном доминировании гетерозиготные особи Aa имеют такой же внешний вид или фенотип, как и гомозиготные по доминантному аллелю АА. Это означает, что для проявления доминантного признака достаточно одного доминантного аллеля, что обозначается как A-.
Если гетерозиготные особи отличаются по фенотипу от гомозиготных и имеют промежуточный фенотип, то говорят о неполном или промежуточном доминировании. Возможным примером неполного доминирования может служить взаимодействие аллелей локуса А.
При кодоминировании у гибридной особи в равной мере проявляются оба родительских признака. По типу кодоминирования наследуется большинство антигенных факторов довольно многочисленных систем групп крови.
В тех случаях, когда анализируют альтернативные признаки, обусловленные парой аллелей одного гена (например, аллели черного B и коричневого b окрасов) говорят о моногибридном скрещивании. Скрещивание особей, отличающихся по двум парам признаков, называется дигибридным, по трем – тригибридным, по многим – полигибридным скрещиванием.
Законы Менделя
Еще в прошлом веке Грегором Менделем были показаны закономерности передачи признаков при скрещивании. Сформулировано три закона Менделя:
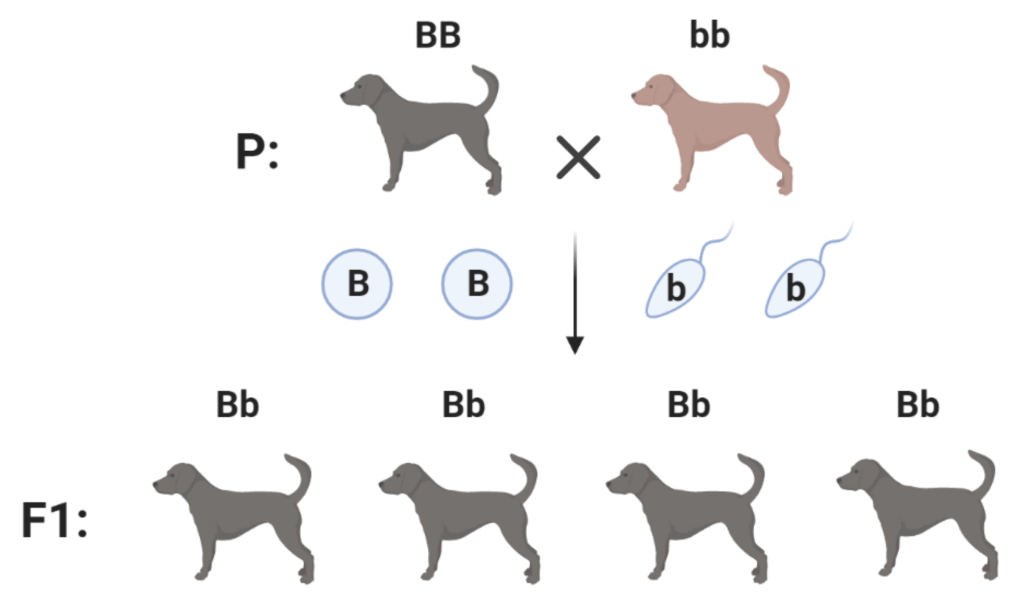
Первый закон Менделя – закон единообразия гибридов первого поколения.
Скрещивание между собой особей, гомозиготных по разным аллелям одного локуса, приводит к рождению гетерозиготных потомков одного фенотипа. Так при скрещивании между собой гомозиготных черных BB и коричневых bb собак все щенки получаются черными Bb.
Особей, относящихся к родительскому поколению обозначают латинской буквой Р, гибридов первого поколения — F1, гибридов второго поколения — F2, гибридов третьего — F3 и т.д.
Второй закон Менделя – закон расщепления гласит: при скрещивании гибридов первого поколения между собой возникает расщепление по фенотипу в соотношении 3:1, а по генотипу 1:2:1. При скрещивании между собой черных гетерозиготных собак с генотипом Bb в помете можно ожидать рождения трех частей черных, состоящих из одной части гомозигот BB и двух частей гетерозигот Bb, и одной части коричневых щенков с генотипом bb.
Мендель сформулировал также правило чистоты гамет, гласящее о том, что гены, находящиеся в гетерозиготном состоянии, не перемешиваются друг с другом, а передаются в половые клетки в неизменном виде.
Определить, кто же из особей с доминантным фенотипом является гомозиготным, а кто гетерозиготным, можно, проведя так называемое анализирующее скрещивание с гомозиготной рецессивной формой. При таком скрещивании в случае гомозиготности исследуемой особи расщепления в потомстве не будет. В случае же гетерозиготности будет наблюдаться расщепление в соотношении 1:1.
Еще одно правило, которое было сформулировано Менделем, носит название правила независимого расщепления аллелей. Оно состоит в том, что во втором поколении каждая пара аллелей и признаков, определяемых ими, ведет себя независимо от других пар аллелей и признаков соответственно.
Для удобства анализа скрещиваний введены графические обозначения, так называемая «решетка Пеннета», в которой в верхнем ряду расписываются гаметы отца, а в левом вертикальном ряду гаметы матери. На пересечении строк и столбцов – генотипы потомков.
В качестве примера приведем скрещивание гетерозиготных черных собак.
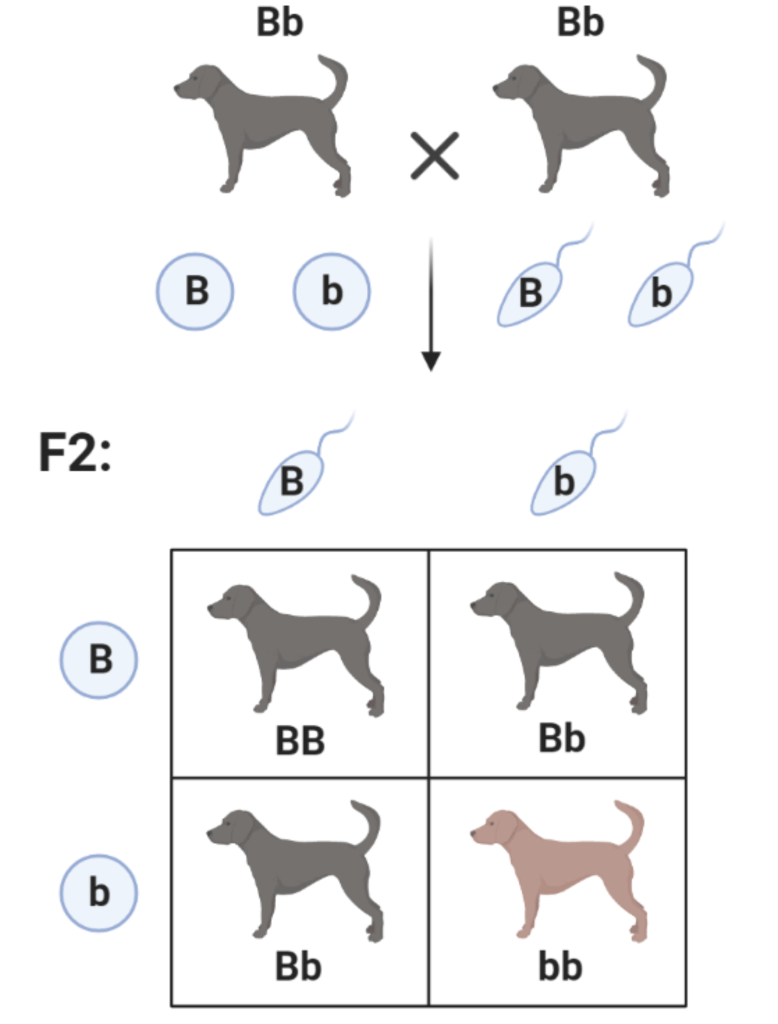
Решетка Пеннета удобна тем, что она автоматически определяет все возможные генотипы и делает удобным их подсчет. В данном случае четко видно, что в потомстве данных производителей произойдет расщепление как по генотипу, так и по фенотипу.
Возможное количество генотипов и фенотипов в потомстве зависит от количества пар анализируемых признаков. Нижеприведенная таблица позволяет определить числовые соотношения в потомстве при полигибридном скрещивании.
| Число пар признаков | Число гамет, образуемых в F1 | Число гамет, образуемых в F2 | Число генотипов в F2 | Число клеток в решетке Пеннета |
|---|---|---|---|---|
| 1 | 2 | 2 | 3 | 4 |
| 2 | 22 = 4 | 22 = 4 | 32 = 9 | 42 = 16 |
| 3 | 23 = 8 | 23= 8 | 33 = 27 | 43 = 64 |
| 4 | 24 = 16 | 24 = 16 | 34 = 81 | 44 = 256 |
| n | 2n | 2n | 3n | 4n |
Качественные и количественные признаки
Все признаки, которыми обладают живые организмы, принято делить на две категории – качественные и количественные. Качественные – это признаки, имеющие четко различимые формы, например, окраска или генетические аномалии, передаваемые отдельными генами. На фенотипическое проявление качественных признаков условия среды практически не влияют. Для характеристики популяции по качественным признакам используют понятия частота генов и генотипов.
Однако большая часть свойств организма представлена количественными признаками. Они проявляют в основном непрерывную изменчивость и могут быть измерены – рост, длина шерсти, вес. Количественные признаки, в большей степени, чем качественные, зависят от условий среды и обусловлены многими генами, так называемыми полигенами, то есть системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называется полимерным. Эти гены также называются аддитивными, так как их действие суммируется. Распределение их численных значений в популяции приближается к кривым нормального распределения. Их наследование можно рассматривать по схеме полигибридного скрещивания.

a — короткоухий кролик, генотип l1l1 l2l2 l3l3; b — длинноухий кролик L1L1 L2L2 L3L3; с — гетерозиготный кролик L1l1 L2l2 L3l3
Селекционеру приходится иметь дело в основном с непрерывной изменчивостью. Менделевский подход к изучению количественных признаков затруднен, хотя они и подчиняются тем же законам классической генетики, что и качественные.
Взаимодействие генов
Гены, находящиеся в разных локусах, также могут влиять друг на друга. При этом можно выделить несколько типов подобного взаимодействия.
Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект действия других генов, называются генами-модификаторами. Изучение окраски у млекопитающих показало, что наряду с крайними формами, обладающими полным развитием пигмента или его отсутствием, наблюдается целый ряд генотипически обусловленных форм. Так, белая пятнистость у собак варьирует от нескольких белых волосков на месте первичной точки депигментации до полностью белой собаки с небольшим пучком окрашенных волос в одном из пигментных центров. В пределах генотипа, определяемого локусом белой пятнистости S, можно выделить массу переходных форм.

Широкую вариабельность, обусловленную генами-модификаторами, имеют чепрачный и тигровый окрасы собак.
Комплементарное взаимодействие генов
Если признак образуется при наличии двух пар неаллельных генов, дающих при совместном действии не тот эффект, который каждый из них обусловливает самостоятельно, то такие аллели обозначают как комплементарные (дополняющие друг друга). В качестве примера комплементарности у собак можно привести взаимодействие генов локусов В и D, определяющих окраску. Локус В отвечает за синтез черного (аллель В) или коричневого (аллель b) пигмента, локус D отвечает за интенсивность эумеланина.
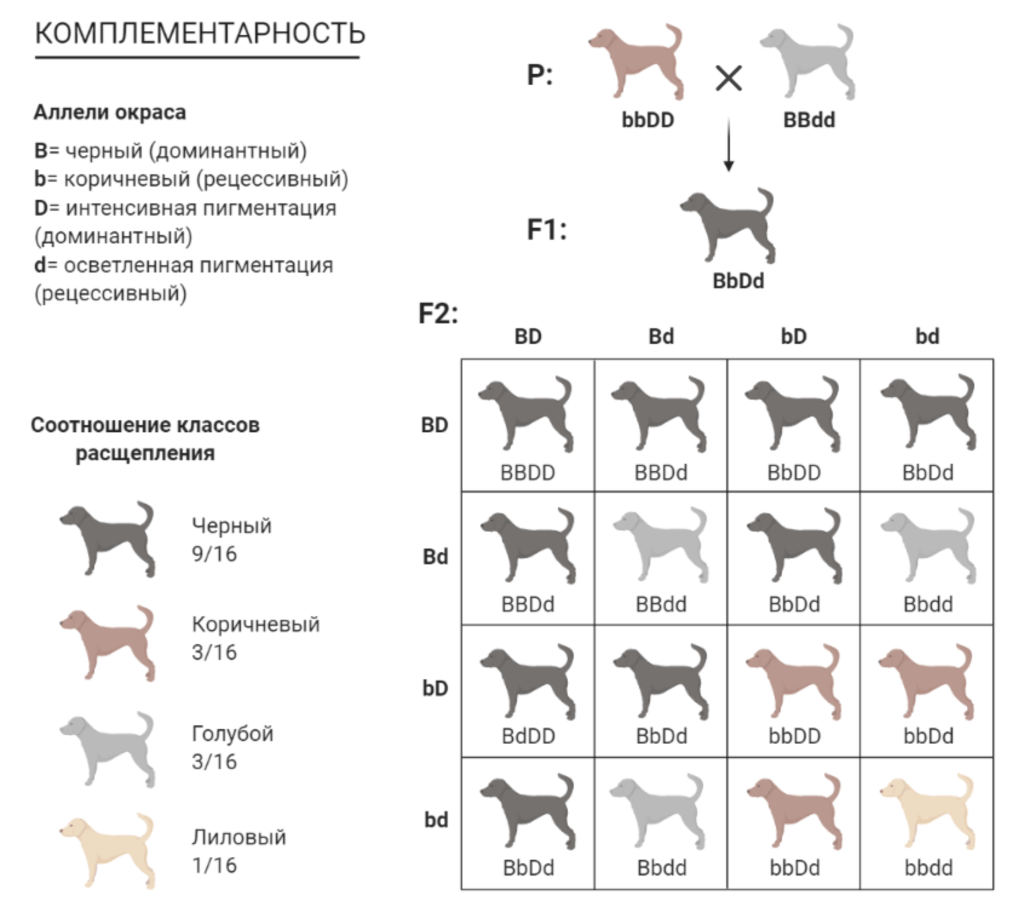
Эпистаз
В любой паре аллелей доминантный аллель препятствует (полностью или частично) проявлению своего рецессивного партнера. Но иногда действие доминантного аллеля подавляется действием гена из другого локуса. Всемогущий ген, который блокирует действие другого гена или генов, называется эпистатичным, а само явление – эпистазом. Гены, действие которых подавлено, называется гипостатичными.
Различают доминантный и рецессивный эпистаз. Примером доминантного эпистаза может служить взаимодействие аллелей локусов К и А, где наличие аллеля доминантного черного КB не позволяет проявляться окрасам агути.
Пример рецессивного эпистаза — взаимодействие локусов B и Е. Формирование окраски собаки зависит от присутствия обоих пар генов. Аллель Е способствует распространению черного или коричневого пигмента по всему телу собаки. Собаки с генотипом ЕЕ или Ее – черные или коричневые в зависимости от аллелей B или b. При Е-ВВ или Е-Bb – собака черная, при E-bb – коричневая. Рецессивный аллель е в гомозиготном состоянии препятствует синтезу в шерстном покрове черного или коричневого пигментов. Соответственно, собака с генотипом ее в окраске шерсти всегда имеет только рыжий пигмент феомеланин.
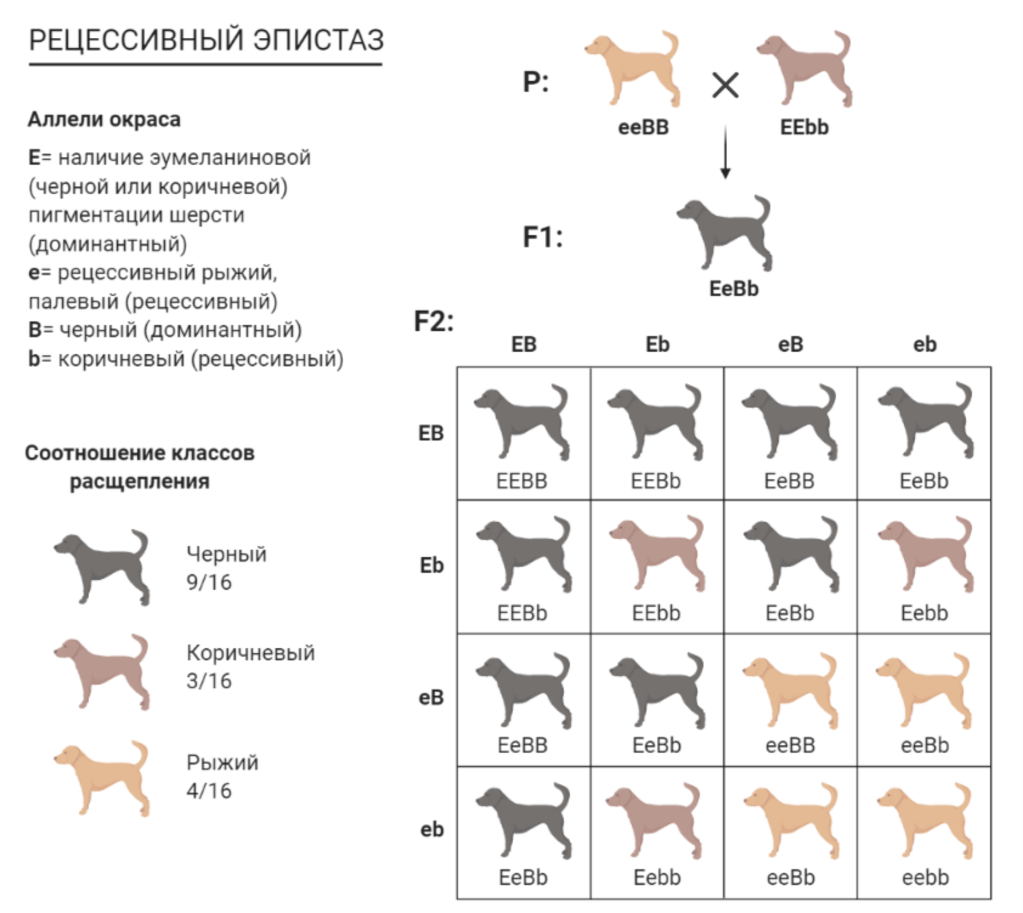
Из рисунка выше мы видим, что в F2 получено 9 черных, 3 коричневых и 4 рыжих собаки. Таким образом наблюдается типичное для рецессивного эпистаза отклонение от теоретически ожидаемого расщепления 9:3:3:1.
По отношению к окраске кожных покровов аллель e не эпистатичен , поэтому цвет мочки носа и обводки глаз у гомозиготы ee будет соответствовать генотипу по локусу В. Собаки с генотипом ееВ- – рыжие с черным носом. Собаки генотипа bbee – обычно палевые или светло-желтые со светлым носом.

черный лабрадор Е-В-; коричневый лабрадор E-bb;
палевый лабрадор с черной мочкой носа и черной обводкой глаз eeB-;
палевый лабрадор с коричневой пигментацией «дадли» eebb
Экспрессивность и пенетрантность
Один и тот же мутантный признак может проявляться у одних и не проявляться у других особей родственной группы. Показатель фенотипического проявления аллеля называется пенетрантностью. Пенетрантность определяется как отношение (обычно в процентах) числа особей, у которых аллель фенотипически виден, к общему числу особей, у которых данный аллель присутствует в необходимом для фенотипического проявления количестве копий (в зависимости от типа наследования может быть нужна одна (аутосомно-доминантный) или две (аутосомно-рецессивный) копии).
Например, если аллель X обладает пенетрантностью 90%, это означает, что из всех особей, у которых данный аллель имеется в необходимом числе копий, лишь у 90% он проявляет себя внешне. При полной пенетрантности (100%) мутантный аллель проявляет свое действие у каждой особи.
У собак достаточно часто встречаются видоизменения хвостов в виде их укороченности, разнообразных изломов и изгибов. Можно предположить, что многообразие этого признака обусловлено его неполной пенетрантностью.
В основе неполной пенетрантности могут лежать как генетические причины, так и влияние внешних условий.
Часто особи, обладающие тем же генотипом в отношении какого-либо наследственного признака, очень сильно различаются по его экспрессивности, то есть степени проявления данного признака. Один и тот же мутантный аллель гена у разных особей в зависимости от влияния генов-модификаторов и внешней среды может проявить себя фенотипически по-разному. Внешняя среда и гены-модификаторы могут изменить экспрессию гена, то есть выражение признака.
В отличие от пенетрантности, которая указывает, у какой доли особей в популяции, генетически имеющих данный признак, он проявится, экспрессивность относится к изменчивости признака у тех особей, у которых он проявился. Так у собак экспрессивность развития прибылых пальцев варьирует от полностью развитых пальцев на обеих задних конечностях до наличия их в зачаточном состоянии только на одной конечности. Подобная вариация экспрессивности характерна и для других наследуемых признаков, в частности и для вышеупомянутых хвостов.
Экспрессивность, как и пенетрантность гена, в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Плейотропия
Достаточно широко распространено явление плейотропии – влияния одного гена на развитие двух и более признаков. Классический «собачий» пример плейотропного влияния – действие фактора Мерля, (локус М окраса). Аллель M в гетерозиготном состоянии Мm дает характерную пятнистость, в то время как в гомозиготном состоянии ММ он ведет к рождению практически белых щенков со значительными нарушениями органов чувств (слух и зрение).

слева направо: первое фото — черно-подпалый окрас, фактор Мерля отсутствует, генотип mm
второе фото — серо-мраморный окрас (blue-merle) шелти, гетерозигота по фактору Мерля (Mm), собака здорова
третье фото — бело-мраморный окрас шелти, гомозигота по фактору Мерля (MM), собака глухая с недоразвитыми глазами (микрофтальмия)
Подобный же пример представляют собой голые собаки. Аллели безволосости в гомозиготном состоянии приводит к внутриутробной гибели его носителей. Все живущие голые собаки являются гетерозиготными и при скрещивании между собой дают в каждом помете гомозиготных особей, имеющих нормальный шерстный покров, так называемых «пуховок».
Явление плейотропии объясняется тем, что гены плейотропного действия контролируют синтез ферментов, участвующих в многочисленных обменных процессах в клетке и в организме в целом и, тем самым, одновременно влияющих на проявление и развитие многих признаков. Но вообще-то говоря, современные исследования показывают, что плейотропным действием в той или иной степени обладают все гены.
Летальные и сублетальные аллели
Некоторые мутации вызывают столь сильные отклонения от нормы, что снижают жизнеспособность организма или даже приводят его к гибели. Такие аллели называют летальными, то есть смертоносными, или сублетальными – снижающими жизнеспособность. В большинстве случаев летальные аллели полностью рецессивны, поэтому гетерозиготные носители этих генов фенотипически совершенно неотличимы от нормальных особей. В гомозиготном состоянии такие аллели могут нарушать нормальный ход развития эмбриона на любых стадиях. О возможности наличия летальных аллелей можно косвенно судить по снижению средней численности пометов или по выпадению некоторой ожидаемой части фенотипов в расщеплении.
Подобный же эффект проявляет и ген куцехвостости Т, который инициирует развитие куцехвостости у ряда пород собак, например, вельш корги пемброк. При этом все куцехвостые собаки гетерозиготны по гену Т, имеют генотип Tt, гомозиготы tt имеют хвосты нормальной длины и формы, а гомозиготы ТТ имеют серьезные отклонения в строении хрящевой ткани, что приводит к гибели щенков еще до рождения.
В случае скрещивания куцехвостых собак между собой вместо ожидаемого соотношения 3:1 получается 2:1, т.е. два куцехвостых и одна длиннохвостая собаки:
Tt х Tt = TT : 2Tt : tt, гдеTT нежизнеспособная собака.
Фактор куцевостости относится к категории доминантных летальных генов, которых значительно меньше, чем рецессивных. При желании его носителей легко изъять из разведения, так как они имеют характерный фенотип.
Летальные аллели одних генов обуславливают крупные аномалии, других – нарушения физиологических процессов. Таких аллелей может существовать сколь угодно много. Показано, что каждый человек является носителем в среднем 4–9 «вредных» или летальных мутаций. Аналогичных результатов можно ждать и по собакам. Известны летальные гены, которые при проявлении в эмбриональном состоянии опасны и для жизни беременной суки, например, при наследственной контрактуре мышц плода, вследствие чего сука не может разродиться.
Признаки, сцепленные с полом
Признаками, сцепленными с полом, называют такие признаки, которые формируются под воздействием генов, локализованных в Х-хромосоме. Наиболее характерным примером наследования признака, сцепленного с полом, является наследование гемофилии собак. У собак, больных гемофилией, в крови не хватает фактора, который, взаимодействуя с кровяными пластинками (тромбоцитами), ускоряет превращение протромбина в тромбин. Гемофилия собак аналогична гемофилии человека и также обусловлена сцепленным с полом рецессивным геном. Ген, ответственный за развитие гемофилии, находится в Х-хромосоме, заболевание имеет рецессивный тип наследования. Следовательно, гемофилия проявляется только у гомозиготных самок (несущих этот ген в обеих Х-хромосомах) и гемизиготных самцов, несущих ген гемофилии в Х-хромосоме. Щенки-гемофилики обычно погибают в раннем возрасте от наружных или внутренних кровотечений. Сохранить до половозрелого состояния такого самца возможно только при постоянном введении специфических препаратов. Самки же неминуемо гибнут не позже первой течки. Гетерозиготные самки внешне абсолютно нормальны и плодовиты, однако половина их детенышей мужского пола страдает гемофилией и половина детенышей-самок оказывается гетерозиготными по этому гену.
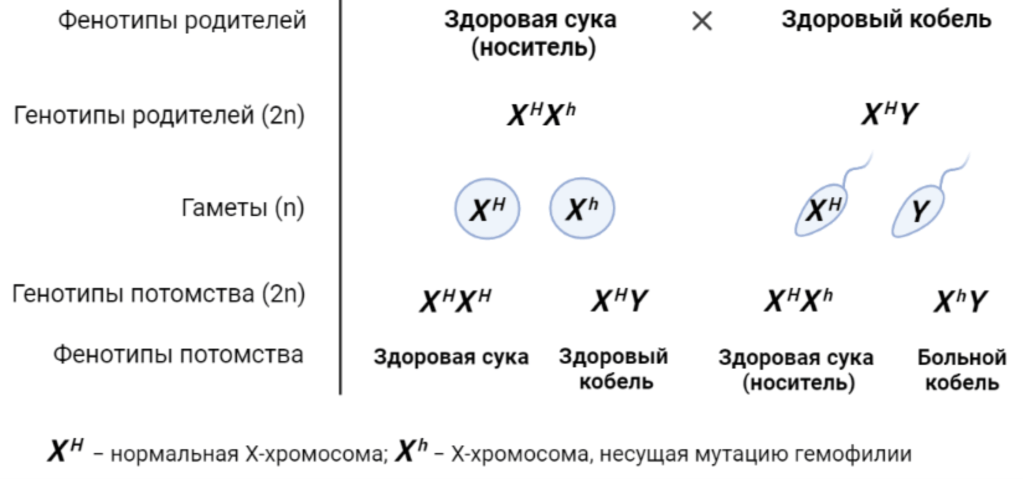
К признакам, сцепленным с полом, относятся также врожденный гипотрихоз, отмеченный у такс и карликовых пуделей; мышечная дистрофия у ретриверов; синдром пошатывания, связанный с гипомиелинизацией и обнаруженный у чау-чау и ряда других пород; подвывих запястья, диафрагмальная грыжа, описанная у золотистых ретриверов и т.д.
Признаки, ограниченные полом
Некоторые признаки совершенно независимо от места локализации генов, их вызывающих, проявляются только у особей одного пола. Это так называемые ограниченные полом признаки. Таковы, например, дефекты развития половой системы, молочность и т.д. Одно из таких явлений – крипторхизм – непрохождение одного или обоих семенников через паховый канал в мошонку. Крипторхизм бывает двусторонний, право- или левосторонний и может быть вызван разными причинами: узостью пахового канала, короткими связками семенников, недоразвитием семенников. Неопустившиеся семенники могут находиться в разных местах брюшной полости. Крипторхизм может быть как врожденным, так и приобретенным. Среди разных его форм существует и генетически обусловленная, однако из-за широкой вариабельности этого признака делать однозначные выводы о его природе невозможно. И уж совершенно неверно трактовать его как моногенный признак, локализованный в Х-хромосоме.
Закон гомологических рядов Н.И. Вавилова
Закон был сформулирован Н.И. Вавиловым в 1920 году. Н.И. Вавилов обнаружил, что все виды и роды генетически близкие между собой характеризуются тождественными рядами наследственной изменчивости. В основе закона гомологических рядов лежит параллелизм генотипической изменчивости у особей со сходным набором генов.
Этот закон имеет универсальный характер. Обнаружено сходство мутаций у разных видов животных. Так были отмечены проявления аналогичных форм аномалий у собак, кошек, кроликов, свиней, человека и др., что указывает на сходство строения многих ферментов и белков и, соответственно, на сходство генотипов. Таким образом, зная формы наследственных изменений у одного вида животных, можно предполагать, что они имеются или могут возникнуть и у другого близкого вида. Признаками, проявляющими гомологию, являются окрас, тип и длина шерсти, вариабельность в размере и т.д.
Ниже приведен пример с гипертрофией мышц у собак и коров. Подобный фенотип описан также для свиней, кроликов, коз и даже кур, встречается у человека, и вызван мутациями в гене миостатина MSTN, который присутствует у позвоночных.


Таким образом, при обнаружении нового фенотипа или аномалии следует поинтересоваться, не описаны ли они для других видов животных. Так, когда у кошек был обнаружен рецессивный рыжий окрас, не сцепленный с полом, первым геном-кандидатом для исследования стал ген локуса E, MC1R, который отвечает за рыжий окрас у большинства видов, и именно в нем в итоге были обнаружены новые аллели. Мутация сиамского окраса собак также была обнаружена в гене, ответственном за аналогичный фенотип у других видов, — гене тирозиназы TYR.
Более подробную информацию по общей и частной генетике собак вы можете получить в книге М.Н.Сотской и Н.Н.Московкиной «Генетика и наследственные болезни собак и кошек» М. 2021. Изд. Аквариум
Для создания некоторых иллюстраций, приведенных в тексте, использовали BioRender.com
